6環境研究
Anatomical study of fruit tree roots
How fruit tree roots respond to environment, especially soil water conditions
- 15 陸の豊かさも守ろう
〈教養教育院〉トゥラダール アスタ(助教)
Astha Tuladhar, College of Liberal Arts and Sciences, Mie University
(This paper is an effort to simplify and present the previous research findings on anatomical changes seen in various fruit tree roots.)
Keywords: exodermis, endodermis, cortex, polyderm, phi-thickening, peridem
Unique features of fruit tree roots
Study of roots especially fruit trees are scarce than other plant parts. However, formation of organizations like the International Society of Root Research (ISRR) and the Japanese Society of Root Research (JSRR) prove that root science is gaining attention as an important area of plant science research. Rootstocks of fruit trees are used for grafting to produce better commercial varieties. Once planted, fruit trees, are hardly transplanted. Therefore, it is interesting to understand how they adapt to local soil environment, including stressful conditions like temporary or long term water-logging and drought. Studying roots that grow deep below ground is challenging due to limited visibility and difficulty to uproot. Also, fruit tree roots become woody as they age. Many questions regarding how woody roots adapt to unfavorable growth conditions remain unanswered today. To understand tree physiology completely studying the adaptive responses of root cells is inevitable.
Role of root tissues and areas of biopolymer accumulation
We hand-sectioned and dyed roots of different fruit trees to observe cross-sections under a fluorescent microscope. Basically, roots provide support and absorb water and minerals. It is said that roots filter the absorbed water at exodermis and endodermis. Cell wall is unique to plant cells. Cell wall of cells that form the exodermis and endodermis accumulate biopolymers like lignin and suberin. Cell wall is considered the first compartment to be affected by stress signals. Biopolymers accumulating in the cell wall are known to play a protective role against fungal diseases and insect attacks. Lignin is a polymer of phenol units that accumulates within the cellulose strands and toughens the cell wall. Suberin, on the other hand, is a polymer with aromatic and aliphatic domains that get impregnated in the intermicrofibrillar spaces of primary cell walls. In our study, we observed biopolymer accumulation in different root tissues like the exodermis, cortex and the endodermis. From previous studies on roots, we know about the Casparian strip at the endodermis. It is a layer of endodermis cells with suberin accumulation in cell walls. It is believed that biopolymer accumulation in woody roots and changes occurring due to stress have not been understood well. So in our study we observed which root tissues accumulate lignin or suberin. Such knowledge may provide further insight to understand their physiological role in plant protection.
What happens to roots when they encounter drought or water-logged conditions?
Studies reveal that anatomical changes occur when roots encounter drought and water-logged conditions. Studying adaptive responses in roots can enhance the selection of appropriate tree species for plantation sites. Identifying how different fruit tree roots respond to soil environment was our purpose. We compared changes in biopolymer accumulation in root tissues among different families of fruit trees. We identified some unique ways of biopolymer accumulation in root tissues like exodermis, cortex and endodermis. Here, we present the anatomical changes observed when roots undergo unfavorable conditions like drought and water-logging. We studied roots of fruit trees belonging to 3 families namely: Mrytaceae, Punicaceae and Rosaseae.
In case of Myrtaceae roots:
In Myrtaceae species, a tissue named polyderm (Fig. 1) formed beyond the endodermis at a distance of 8-10 cm from the root tip, when the outer surface of the root changed from white to brown. Polyderm is a unique tissue with alternative layers of cells accumulating lignin and suberin. The lignin accumulating cell layers were called thick-walled cells (twc) and suberin accumulating cell layers that also contained the Casparian strip were called enderdermis-like cells (elc). We found deposition of lignin and suberin that form a unique pattern in five species in the Myrtaceae family namely: 1) rose apple (Syzygium jambos), 2) strawberry guava (Psidium cattleianum Sabine), 3) wax apple (Syzygium samarangense), 4) guava (Psidium Guavaja), 5) feijoa (Feijoa sellowiana) wax apple.
We observed that biopolymers that deposit in the cell wall, causing it to thicken, are site-specific. Lignin accumulating in the exodermis was “m”shaped, however, lignin accumulating in cortex cells and thick-walled cells were “w”shaped and this was consistent in all five species of Myrtaceae. It is not yet understood what factors determine such site specificity of biopolymer deposition in plant root tissues (Tuladhar et. al, 2014).
In wax apple roots (Myrtaceae), the number of polyderm layers that formed beyond the endodermis was influenced by soil water availability. Under flooded conditions the number of polyderm layers was fewer than roots grown in normal conditions (Tuladhar et.al, 2015). Decrease in polyderm layers means there were fewer cell layers with lignin and suberin indicating that the role of lignin and suberin might be greater under drought conditions.
In feijoa roots (Myrtaceae), the timing of lignin accumulation in thick-walled cells was observed. Lignin accumulation was earlier in roots grown in drought conditions than in normal conditions (Tuladhar et. al, 2016). From these observations, it was confirmed that lignin accumulation in tissues increased and its accumulation became earlier when encountering drought stress (Fig. 2).
In case of Punicaceae:
In pomegranate roots (Punicaeae) it was revealed that its roots have lignin accumulating periderm(Fig.3).Unlike Myrtaceae species, alternative pattern of lignin and suberin accumulating cell layers was absent in this species. However, a periderm, several cell layers deep, existed beyond the endodermis. The cell layers forming the periderm accumulated both lignin and suberin in the same layer instead of forming separate layers for lignin and suberin (Tuladhar et. al, 2014). Similar observation of periderm was also conducted in grapevine roots but lignin accumulation in periderm was unique to Punicacaeae species.
In case of Rosaceae:
In loquat roots (Rosaceae), we found that a cell-wall ingrowth caused by the deposition of lignin in the shape of a Greek letter phi is commonly found in cortex cells adjacent to the endodermis. Other than endodermis and exodermis which are important for plant resistance, cortex cell wall thickenings should also be studied because it is a unique and flexible tissue found only in new roots undergoing primary growth. For perennial woody fruit trees, cortex cell wall thickening may be providing mechanical support to roots but they may also be functioning as defense structures like the endodermis and exodermis. So far, phi-thickenings have not been confirmed in seed germinated loquat roots. Development of phi-thickening in water-logged condition was studied for the first time. Our studies confirmed that phi-thickenings were present in loquat roots grown in water-logged conditions as well though the number of cortex cell layers forming phi-thickening was fewer compared to drought conditions. Suberin-like accumulation was also observed in the intercellular space of exodermal cells and the number of cell layers with suberin deposit depended on water availability (Fig. 4).
Conclusion:
Biopolymer accumulation causes structural changes in root tissues. Its accumulation increases under drought conditions but decreases under water-logged conditions. The Casparian strip, polyderm, periderm, phi-thickening are all names of different root tissues accumulating biopolymer (such as lignin and suberin). The function of each is somehow related to protection against diseases, insect attack or stressful growth conditions. The nature of accumulation may vary among fruit tree species, for example, the polyderm is a unique tissue found only in Myrtaceae, Hypericaceae, Onagraceae and the occurrence of phi-thickening is consistent in Rosaceae and a periderm with lignin and suberin accumulation in the same cell layer was unique in Punicaeae.
From our initial studies on fruit trees, it has become clear that phenolic and aliphatic compounds, such as lignin and suberin, deposition on cell walls are influenced by root age and soil water availability. Recent studies confirm that deposition of biopolymers in the cell wall is highly site-specific. For example, suberin deposition in the epidermis, exodermis and endodermis plays an important defensive role in root function and protects against stress.Roots can exhibit significant difference in the development of endodermal and exodermal layers among species.
How do growth conditions affect biopolymer accumulation in a specific root tissues? Endodermal Casparian strip was present closer to the root tip in drought conditions. Similarly, lignin accumulation in the form of phi-thickenings increased dramatically in loquat roots grown under drought conditions. Thus, in this study, variation in deposition of lignin and suberin in specific root tissues was studied in various fruit tree species according to root age and growth conditions.
References:
- Tuladhar, A., Ohtsuka, S., & Nii, N. (2014). Formation of exclusive pattern during accumulation of ligno-suberic material in cell wall of Myrtaceae root tissues including epidermis, exodermis, endodermis and polyderm. Plant Root, 8, 24-32.
- Tuladhar, A., & Nii, N. (2014). An anatomical study of developmental changes in maturing root tissues of pomegranate (Punica granatum L.) and formation of a unique type of periderm. Plant Root, 8, 33-41.
- Tuladhar, A., Ohtsuka, S., & Nii, N. (2015). Anatomical study on wax apple (syzygium samarangense) roots under long-term water-logged conditions. Tropical Agriculture and Development, 59(1), 1-6.
- Tuladhar, A., Ohtsuka, S., & Nii, N. (2016). Effect of Soil Water Conditions on Biopolymer Accumulation in Root Polyderm of Feijoa sellowiana (Myrtaceae). Tropical Agriculture and Development, 60(3), 195-199.
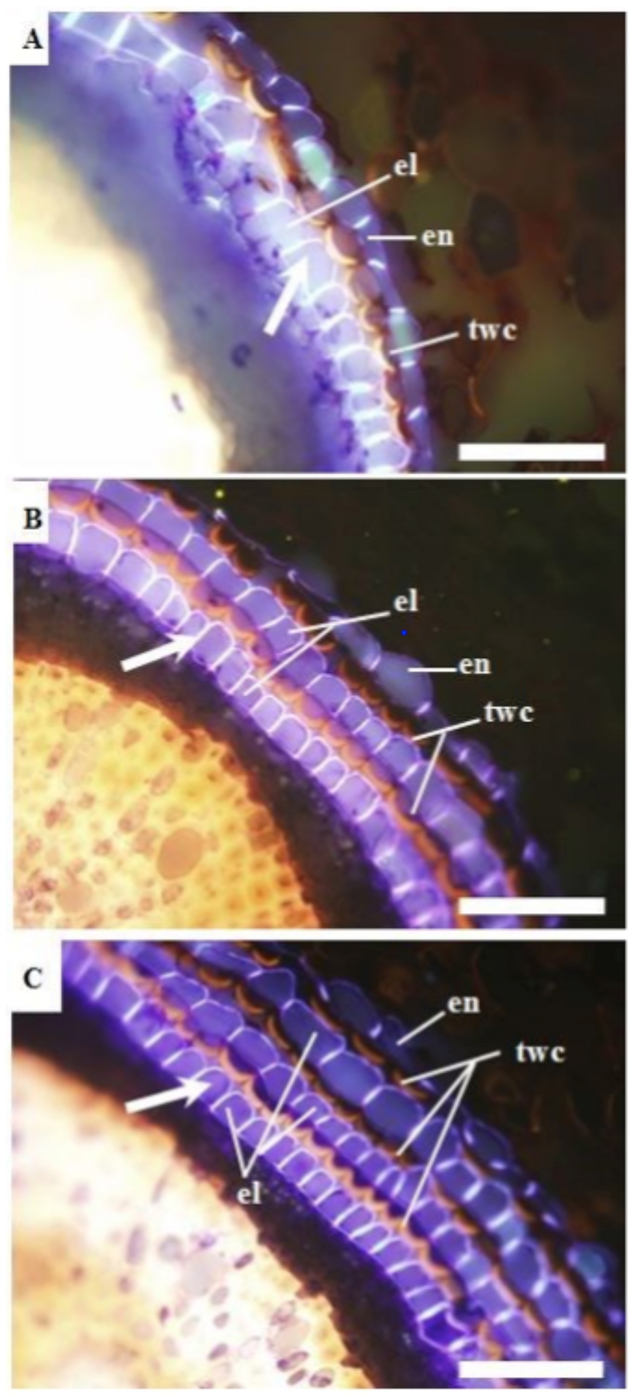
【Fig.1.】 Formation of a unique alternating pattern of polyderm with “m”shaped suberin layers and “w”shaped lignin layers that increased under drought stress conditions in Myrtaceae roots. en: endodermis, Cs: Casparian strip, twc: thick-walled cells, el: endodermis-like cells. Scale bar=50 μm.
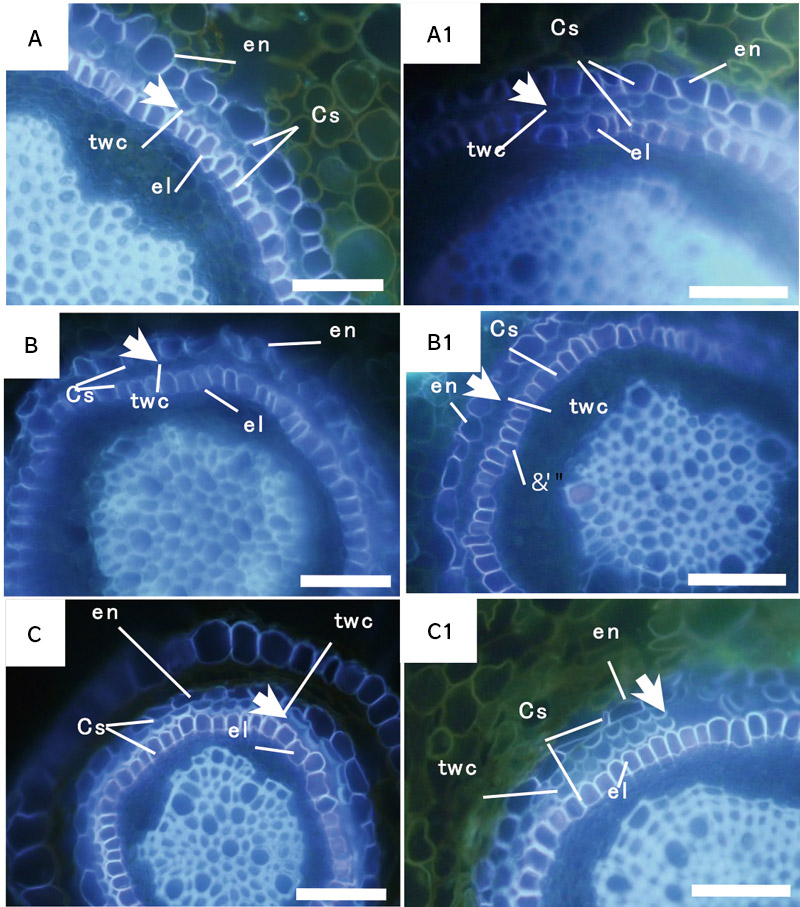
【Fig.2.】 Difference in biopolymer accumulation in thick-walled cells (twc) in feiijoa (Myrtaceae) roots grown in normal (A, A1,), water-logged (B, B1), drought (C, C1) treatments. Arrowhead shows the extent of biopolymer accumulation in thick-walled cells. en: endodermis, Cs: Casparian strip, twc: thick-walled cells, el: endodermis-like cells. Scale bar=50 μm.
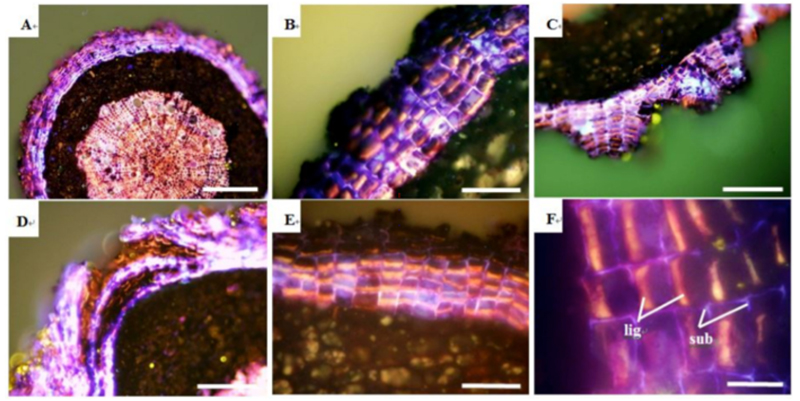
【Fig. 3.】 Cross-sections of pomegranate (Punicaceae) roots showing the formation of lignified cell layers at the periderm. Outermost layer (A) in woody roots after endodermis, cortex, exodermis shed off. Enlargement of (A) is (B). Triangular chunks of lignified cells at with opening in the middle (C). Lenticel-like increase in thick-walled lignified cells at the root surface (D). Lignin-like accumulation which is “closed” centripetally and suberin-like accumulation in the cell wall resembling a network (E). (F) is enlargement of (E). lig: lignin-like substances accumulating layer, sub: suberin-like substances accumulating layer. Scale bar=25μm. (F), 50μm (B, C, E), 100μm (D) 500μm (A).
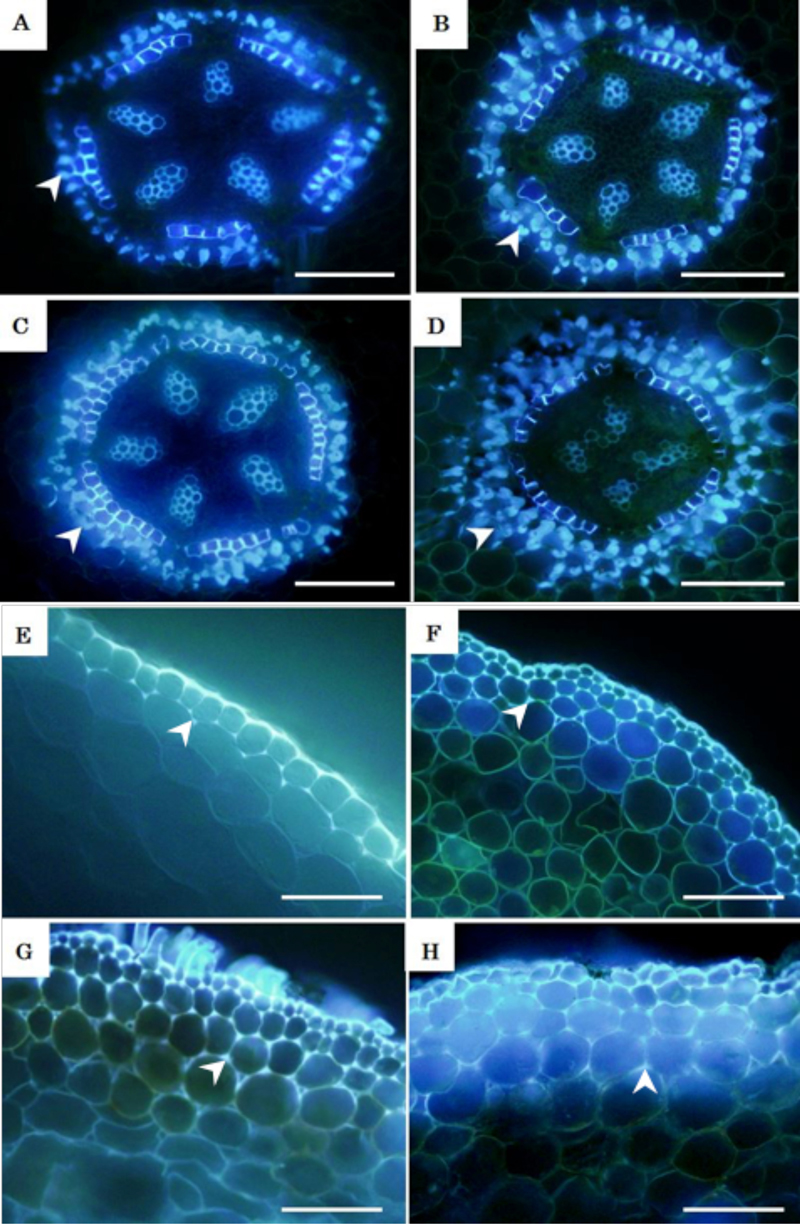
【Fig.4.】 Extent of phi-thickening (arrows in A-E) and extracellular suberin-like substances accumulation or ESA (arrows in E-H) in loquat (Rosaceae) roots. A, E represent roots under water-logged conditions. B, F are roots grown under normal conditions. C, G are roots grown in drought conditions. D, H are roots grown in extreme drought conditions. Scale bars=50 µm (A- H).